PDF of this article (293 KB)



Wendy Nelson and Anne-Maree Schwarz report on the sex lives of large brown algae.
Around the coasts of New Zealand, forests of brown seaweeds fringe the rocky shores and reefs. Although often called kelps, only three of the native large brown algae found around our shores are true kelps or laminarians (classified in the order Laminariales). These are Macrocystis pyrifera (giant kelp), Ecklonia radiata (common kelp), and Lessonia variegata. The rest of our large brown algae are either bull kelps (4 species) or fucalean algae (28 species).
All of these large brown algae are important ecologically, giving three-dimensional structure and complexity to rocky reefs, and functioning like terrestrial forests. They form canopies that provide shelter, food, and habitat for many other species – algae, invertebrates, and fishes. Around the world, kelp species are extensively harvested from wild populations and some species are farmed in large-scale marine farms. Kelps are used for human food, animal and stock feeds, as well as fertilisers and soil conditioners, and as a source of many useful extracts, some with human health benefits and others employed in industrial processes.
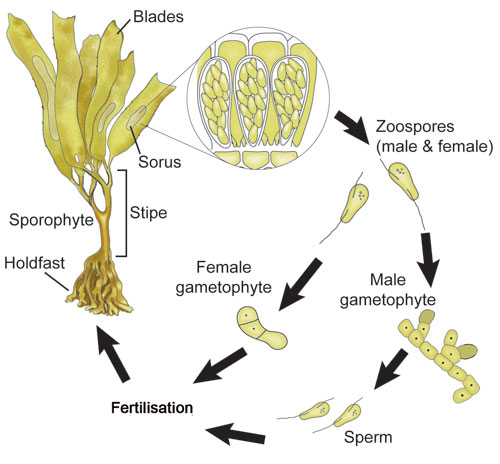
The bull kelps and fucalean algae have 'direct' life cycles and reproduce via eggs and sperm. However, the true kelps, or laminarian algae, have a different approach to reproduction: they have 'heteromorphic' life cycles (hetero = different, morph = shape/appearance), and alternate between two very different forms. Large kelps that you see on the shoreline are the ’sporophyte' (spore-bearing) phase, and these alternate with a microscopic 'gametophyte' phase. Gametophytes are so small they have rarely been found in the wild; it is presumed they live in cracks and crevices in rocky reefs. Almost everything we know about kelp gametophytes has been discovered from laboratory studies.
Why study seaweed life cycles?
Recently NIWA staff have been studying Lessonia variegata, one of our true kelps, which is found only on New Zealand’s wave-exposed coasts. Despite its widespread distribution around New Zealand, very little is known about L. variegata, including its growth and reproduction. There is commercial interest in extracting compounds found in the blades of Lessonia, and also in using it to feed farmed paua. Species of Lessonia in Chile are extensively harvested, and research there is exploring farming and enhancement techniques. In our research, we investigated the life history of L. variegata as part of a wider study of the ecology and reproductive cycle of this species.
Why is it important to understand life cycles in seaweeds? By understanding the reproduction of the species we can gain important insights about how species interact with environmental conditions. The way in which these large brown algae reproduce has a profound effect on their ecology.
Seaweed sex at the beach
From field experiments on the Wellington south coast we found that Lessonia was fertile from autumn through to spring, with fertility peaking in midwinter. Dark-brown patches called sori (singular = sorus) develop on the sporophyte blades (see diagram). If you run your hand along the surface of a fertile blade, you can feel the raised surface and the velvety texture of the sorus. Within the sori there are closely packed ’sporangia' filled with many spores. By November, almost no sori could be found on the blades of Lessonia. The end of the fertile period coincided with the onset of longer days and the warmer temperatures of summer.
When the zoospores are released, they swim and then settle, germinating to form microscopic male and female gametophytes. The male and female gametophytes produce sperm and eggs; the sperm can swim but the eggs do not move. As these gametes are formed on separate individuals, the sperm must be able to locate a female gametophyte in order to fertilise an egg. The kelps have evolved chemical attractants: the females release compounds that attract the swimming sperm. These chemical attractants can operate only over quite small distances in seawater. Once fertilisation has occurred a new sporophyte kelp begins to develop.
Seaweed sex in the lab
In the lab we grew gametophytes of Lessonia under different conditions, varying the temperature, daylength, and light intensity. In these culture experiments, in 14 days the male and female gametophytes had germinated from zoospores, formed egg and sperm cells, and the sperm had successfully located and fertilised eggs. More gametes were formed in warmer (15 °C) rather than colder (10 °C) conditions – and under longer rather than shorter days (15 vs 9 hours). Under low light conditions the gametophytes didn't form sperm or eggs but instead grew as filamentous clumps. When the gametophytes were moved from winter conditions to summer conditions, or from low to higher light, they responded rapidly, forming gametes and developing larger sporophytes under summer growth regimes. This indicates that the gametophytes are able to respond swiftly to shifts in environmental conditions that would be typically experienced in late winter/early spring.
People working on other kelp species have also observed that filamentous growth in gametophytes occurs when the conditions are not ideal. They can survive in this state when the environmental conditions are not suitable for further development – although they may have enough energy to grow, the gametophytes require certain minimum conditions to form gametes. The gametophytes serve as a 'seed bank' for the kelp forests: when the seawater conditions are 'right' then the gametophytes switch to producing eggs and sperm.
More lessons to be learned from Lessonia
In our research we have looked at Lessonia from central New Zealand in relation to temperature and photoperiod conditions experienced in this part of the country. We are still at an early stage in understanding the ecology of this native kelp. Do Stewart Island Lessonia variegata respond in the same way as individuals from Wellington or from the Poor Knights Islands? Are there particular conditions under which Lessonia cannot grow? Which stage is more vulnerable, the gametophyte or the developing sporophyte? In Chile, populations of a species of Lessonia were negatively affected by El Niño conditions in the 1980s and have not recovered. Kelp species are known to be vulnerable to various kinds of human-mediated changes and pollution sources – increased sedimentation, sewage outfalls, mine tailings. In Southern California, the abundance of giant kelp has closely mirrored long-term fluctuations in oceanic conditions, and may prove to be an indicator of ocean warming.
To manage kelp populations sustainably we need to understand the demography of the species – the numbers of new individuals arriving in a population and their subsequent growth, reproduction, and death. Having a good understanding of the life history of a species is a fundamental part of building such a picture.
Sussing out seaweeds
- Of New Zealand’s many species of large brown algae, only a few are true kelps.
- Large brown algae are important ecologically and have economic value.
- True kelps, including Lessonia variegata, have two phases: a large, bladed form and a microscopic stage.
- Our lab studies have helped determine the optimum environmental conditions for kelp reproduction.
Dr Wendy Nelson works on various aspects of New Zealand marine macroalgae: taxonomy, life histories, and ecology. She is based at NIWA in Wellington. Dr Anne-Maree Schwarz is a benthic ecologist with interests that range from seaweeds to seagrasses and from the tropics to Antarctica. She has recently left NIWA to work for VSA in the Solomon Islands.
Author contact: [email protected]
Teachers’ resource for NCEA AS: Biology 90460 (2.4), 90463 (2.7), 90717 (3.5); Science 90772 (2.3). See other curriculum connections at www.niwa.co.nz/pubs/wa/resources