It’s now almost ten years since blooms of the stalked diatom Didymosphenia geminata – commonly known as didymo, or “rock snot” – were first discovered in the Waiau River, Southland. Not only did the discovery trigger a major biosecurity response here in New Zealand, but it also highlighted didymo as a potential threat to rivers worldwide.
Didymo was known to have a wide distribution in parts of the Northern Hemisphere, with historical reports of blooms in some countries (e.g., Norway and the UK). However, the New Zealand discovery led to a general belief that we were dealing with an aggressive invasive organism being spread by humans. Certainly the pattern of spread of didymo in the South Island of New Zealand was consistent with that view. At the same time, no didymo has yet been detected in any North Island rivers, despite ongoing surveillance.
Didymo blooms are unusual because they typically occur in rivers with low nutrient concentrations, i.e., low supplies of nitrogen (N) and phosphorus (P); algal blooms are usually linked to high nutrient concentrations. In New Zealand, this was a particular concern because it seemed that some of our most iconic and pristine rivers were at risk. In suitable conditions, didymo produces thick smothering mats covering large proportions of the river bed. The mats (comprising mostly a polysaccharide stalk material) can lead to changes in communities of invertebrates and other algae on the river bed.
The blooms look unsightly and are a nuisance. They affect river users, interfering with angling, sporting and recreational activities, and blocking water intakes.
At the time of the NZ discovery only limited information was available on didymo. Since then there has been considerable research on various aspects of the distribution, ecology, and effects of the species.
A significant breakthrough in understanding didymo resulted from of a series of experiments and surveys in New Zealand. Three findings were pivotal.
First, we demonstrated phosphorus (P)-limitation of didymo cell division rates in water from a river with large blooms. In other words, adding P (as dissolved reactive phosphorus, DRP) caused cell division rates to increase, even though large blooms were already present.
Second, we showed that the polysaccharide stalks of didymo (which form much of the bloom material) grew longer when DRP concentrations were low and light was high.
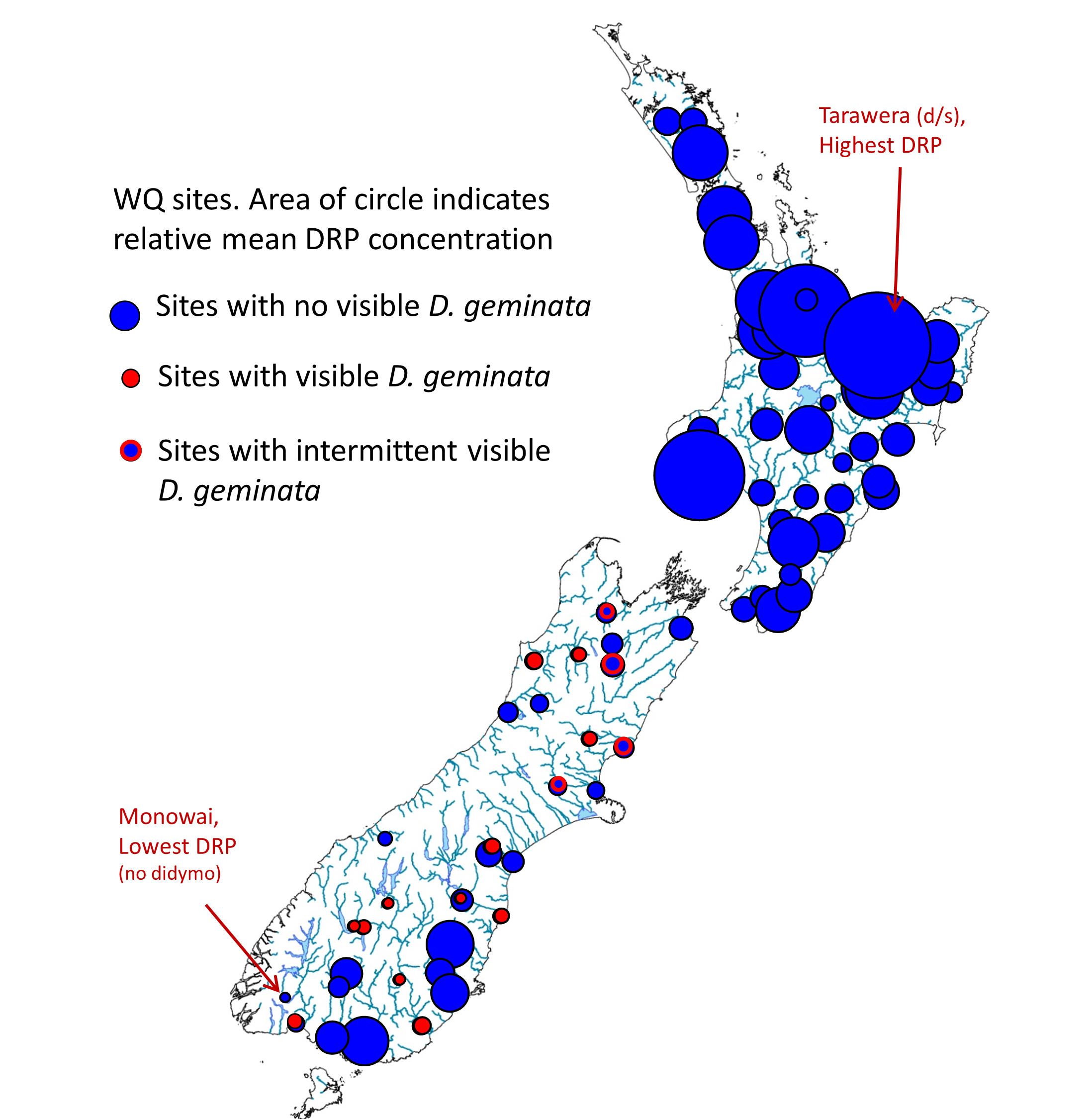
Third, we showed that low DRP was associated with low cell division rates and high didymo cover in rivers across the South Island: visible didymo was not observed at sites when long-term DRP concentrations (e.g., 1 – 2 year average) were more than about 2 ppb.
All this led to the conclusion that the blooms are caused by low P concentrations. The mechanism is that when DRP levels are very low, the energy that would have been used to make new cells is diverted into production of carbohydrate stalks – it is the stalks that make up the bulk of the blooms. This realisation that blooms occurred because of low P solved a puzzle that had, until then, been approached from another angle: that didymo was able to form blooms in low P conditions through some mechanism of acquiring additional P.
Low P as the cause of blooms provides answers to a few questions about distribution. First, taking a New Zealand perspective, why has didymo remained undetected in the North Island? Quite simply, dissolved P concentrations in North Island rivers are too high.
Many North Island rivers have average DRP concentrations greater than 2 ppb, as a result of P-rich catchment geology, more intensive catchment development than in the South Island, or both.
A recent analysis of data from the National River Water Quality Monitoring Network (NRWQN) confirmed the clear difference between islands. It’s less clear why not even a single live didymo cell has been detected in any North Island river. After 10 years, even with efforts to prevent transfer via humans, it is unlikely that no cells have been introduced. It has been shown in experiments that water chemistry, including high P concentrations, doesn’t prevent initial attachment and growth (12, 13). Therefore short-term survival might be expected. However, unless concentrations of P decline significantly (to levels below 2 ppb), blooms will not form in most North Island rivers.
Second, taking a global perspective, what caused didymo blooms to suddenly appear in some regions of the Northern Hemisphere in the early 1990s?
The low P – didymo bloom link supports a plausible hypothesis. Declines in P concentrations have been documented in some northern rivers, and a range of processes may lead to such declines, from local scale (e.g., river impoundment) to regional-continental scale (climate change) (10). There are historical records of didymo presence (but as a rare species) in many areas which are now experiencing didymo blooms. If conditions became suitable (i.e., P declining to below 2 ppb, on average), then the blooms would follow (10).
At this stage, to our knowledge, there are still no data that clearly link the appearance of any recent didymo blooms in the Northern Hemisphere to declines in dissolved P. But back in New Zealand, data from the NRWQN do not show any general decline in DRP in South Island rivers in the years leading up to the sudden appearance of blooms in 2004. We have no doubt that didymo is a recent introduction. There are no historical records of didymo in the South Island, and only one (highly likely a case of mistaken identity) from the North Island (3).
A likely scenario is that didymo arrived in New Zealand following the transformation of didymo in the Northern Hemisphere from a rare species with only occasional blooms in some locations to a common species with widespread blooms. In an age of increasing tourism and air travel, the transfer of cells to remote locations like the South Island – which also happened to have rivers with perfect conditions for blooms – was almost inevitable.