NIWA expends considerable effort on determining the age of commercial fish species. But why age a fish? And how do we do it?
Overview
Why age a fish?
Age information contributes to the monitoring and management of fishery resources in New Zealand by enabling:
- calculation of growth rates. By ageing numerous individuals over the full range of sizes in a population, it is possible to generate a growth curve (i.e. average length at each age) for that population. This enables us to determine how productive the species is and what variability there is in growth rates over time
- calculation of mortality rates. By ageing a random sample of individuals from a population, and noting how the frequency of fish in each year class (all of the fish born in the same year, otherwise known as a cohort)declines with increasing age, it is possible to estimate the rate of natural mortality (if the sample is from an essentially unfished population), or the combined rate of mortality attributable to natural causes and fishing (if the population has been significantly fished).
- estimation of population age structures. The aged random sample described above also shows what age classes the population is composed of, or, if the sample is from a particular fishery, what group of age classes that fishery exploits.
- estimation of annual spawning success. An aged random sample can indicate the relative strengths of individual year classes (after accounting for mortality over time), and show how successful spawning was in each year. The more estimates that can be derived for the strength of a particular year class (e.g. from consistent sampling in consecutive years), the greater the confidence in the estimate for that year class. These estimates of year class strength can explain variations in total stock abundance and be used to predict future recruitment to the fishery.
- investigations of stock structure. Numerous fish species in New Zealand waters are represented by two or more stocks (i.e. distinct populations of the same species with little or no genetic or physical mixing). Differences in growth rates or population age structures between areas can indicate (but do not prove) that multiple stocks occur.
Approach
How are fish aged?
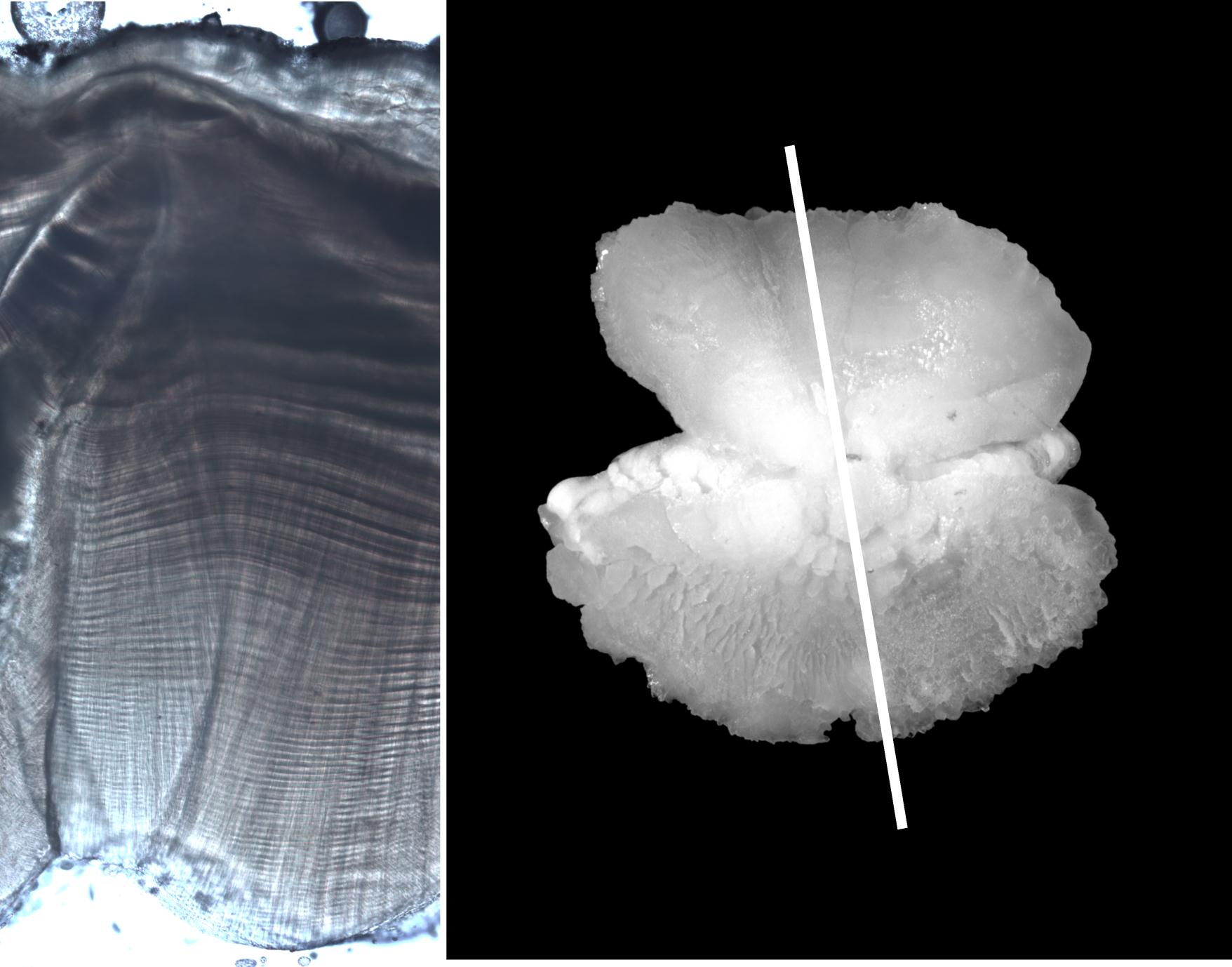
In the plant kingdom, tree rings indicate annual growth increments. In the animal kingdom, probably the most well-known patterns visible externally are on the shells of molluscs and tortoises. However, other groups have also been shown to produce annual or daily growth increments: these include some coral, cricket and starfish skeletons, and the teeth of some mammals. However, fish produce growth increments in various calcified structures, most notably in their otoliths (or 'ear stones'), but also in scales, spines, vertebrae and other bones.
Otoliths are paired calcified structures occurring in the heads of all bony fishes - they do not occur in the cartilaginous sharks or rays. Like the human ear drum, otoliths are part of the fish's sensory and balance system, and the name 'otolith' comes from two Greek words meaning ear and stone. Otoliths are mainly calcium carbonate (limestone) with a small component of non-organic trace elements and protein molecules.
It is estimated that well over a million fish are aged each year worldwide, highlighting the importance of age information in fisheries science. Fish ageing studies are either based on annual or daily growth. Annual ageing studies are the most common because they provide information on fish productivity and population structures. Daily ageing, using otolith microstructure, is used mainly in studies of young fish.
Poster: validating an ageing method for the Antarctic toothfish (PDF 16 MB)
All ageing investigations involve a count of microscopic contrasting light and dark zones, though methods vary between species and structures. Scales and some otoliths are examined whole; other structures (i.e. most otoliths, spines and bones) are sliced transversely (like cutting through a tree trunk) and the zones are counted across the cut surface. The sliced structure may be untreated, or baked or stained to enhance the visibility of the contrasting zonation pattern. Vertebrae are sectioned or x-rayed, showing regular patterns of higher and lower density.
Unfortunately, the growth increments formed in fish parts are not as clear as tree rings; if they were, the studies of fish population dynamics would be much simpler and more precise than they currently are. There are three major problems encountered during fish ageing:
- not all structures in a fish show a complete growth record throughout the fish's lifetime. For example, scales may show a clear pattern of growth increments when the fish is relatively young and growth is rapid, but the increments may then merge at older ages when growth in size becomes negligible.
- the interpretation of the growth zones can be difficult: zones can be diffuse, variable in width, and have confusing microstructures.
- finally, it is essential to know how often new zones are made (their 'periodicity'), and this can be a difficult process.
Because ageing information contributes in so many ways to population modelling and management, any inaccuracies in age data can result in management actions which could impact seriously on the population being fished. There are examples worldwide where ageing error (usually underestimation of age) has contributed to the serious overexploitation of a fish population: New Zealand examples are orange roughy and bluenose.
We now know that these species are at least twice as old as they were thought to be when they were first fished. When the orange roughy fishery began in the early 1980s the longevity of this species was believed to be about 30 years, and it was subject to very heavy fishing. However by the end of the 1990s, it had become apparent that orange roughy had very low productivity levels, could live to at least 100 years, and could not sustain the previously high exploitation levels.
Outcomes
The current status of New Zealand fish ageing
Before 1990, even though age estimates or growth information was available for 62 New Zealand species, including about 45 commercial species, published growth parameters (i.e. the relationship between length and age) were available for only 15 commercial species. The ageing technique had been comprehensively validated for 4 of these 15 species only, though partial validation had been achieved for most others (generally for the juvenile part of the growth curve).
Many of New Zealand's commercial species grow quite slowly and to relatively old ages, so their otoliths are difficult to interpret. The complex way in which zones are structured in some otoliths has been partially attributed to the relatively deep waters that New Zealand species inhabit, where seasonal influences on the environment (and hence, on growth) are more blurred than in shallow, coastal waters.
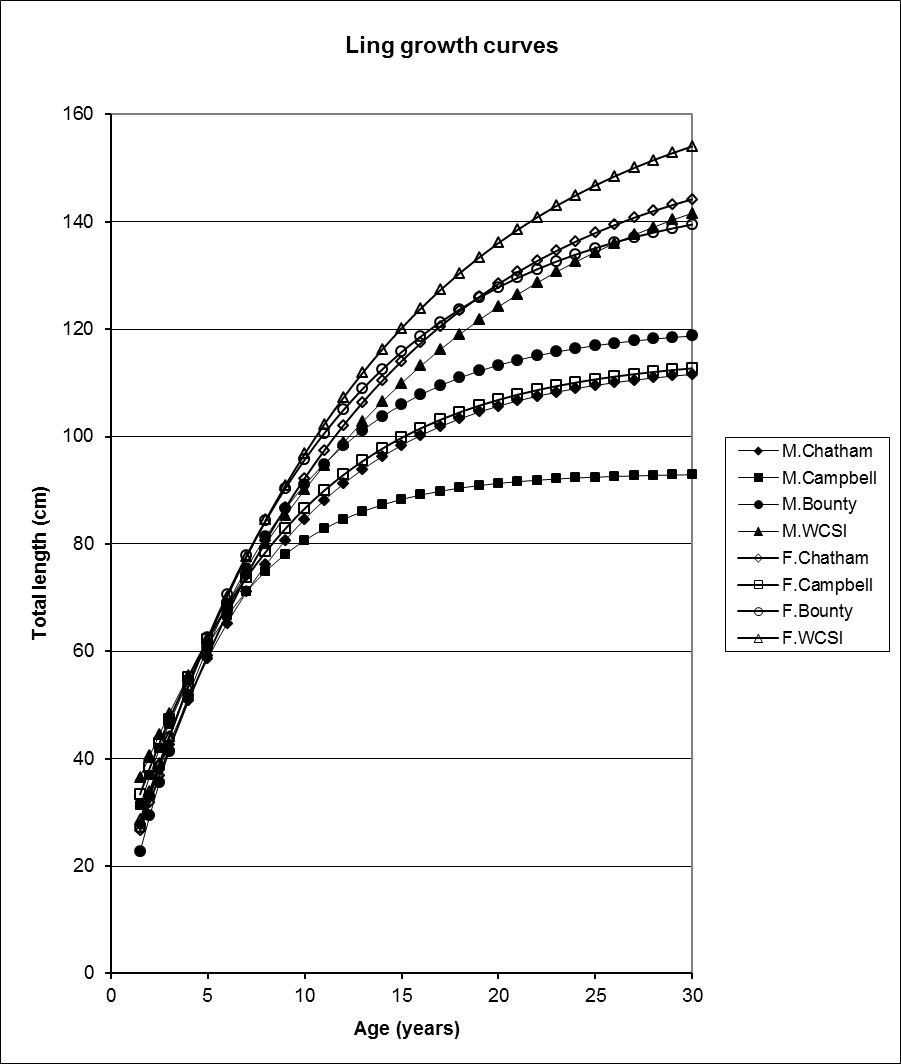
Since 1990, considerable progress has been made in ageing New Zealand fishes. Growth curves are available for about 60 commercial species and about 40 of these are considered fully or partially validated. Usually a single growth curve is not sufficient to describe growth patterns for a single species: growth often differs between sexes, with females generally growing faster and to a larger size than males. Also, many species comprise distinct biological stocks (i.e. geographically and/or genetically distinct) and growth can vary between stocks. For example, there are at least five stocks of ling in New Zealand waters, each with significantly different growth rates.
Several different methods for validating the periodicity of otolith growth zones have been successfully used for New Zealand species. Snapper is an example of a species with a comprehensively validated ageing method. Over 4600 fish covering a wide range of sizes (and ages) were caught, marked with an external numbered tag, measured, injected with oxytetracycline, and released. Over the following 3 years, 1113 of these fish were recaptured and their ages at death estimated from counts of zones in sectioned otoliths.
Oxytetracycline (an antibiotic) deposited in bones and otoliths at the time of injection leaves a layer that is visible (fluoresces) under UV light. It is clearly visible in sectioned otoliths, and if the number of dark zones in the otolith formed outside the fluorescent band is the same as the number of years since injection, this shows us the annual formation of zones.
This was the case for snapper. Additional information from the study further strengthened the validation of the ageing method. Lengths at the times of tagging and recapture were known for all fish, so these data could corroborate the growth curve calculated from the otolith counts.
The validation of the ageing method for hoki, New Zealand's largest commercial fishery, was achieved using two methods. The juvenile growth section of the otolith is difficult to interpret because the zones are often diffuse, with complex micro-banding. However, the growth of hoki in its first three years is fast enough to produce distinct groups (called modes) of similar-sized fish with each mode representing a different year class. So, by examining otoliths from fish in these distinct modes, it was possible to decipher and validate the early growth patterns in the otoliths.
Validation of annual zones in otoliths for ages four to eight years was achieved by following the progression of strong year classes in population age structures (i.e. the number of fish in each year class) calculated in each of seven consecutive years. Similar patterns of strong and weak year classes were identified in each distribution (i.e. fish 'born' in 1983, 1984, and 1987 were always abundant, whereas those from 1986 were rare), showing that the counted zones were formed annually.
One of the more complicated age validation studies is of bluenose, a species which spends its juvenile life in surface waters, but lives in relatively deep waters after about three years. Bluenose otoliths have a very complex structure, with two possible interpretations. Initial growth is clearly rapid, but after about three years the zones can be interpreted as either abundant and narrow, or relatively broad but composed of complex microstructure. As a result, maximum age was estimated to be about 12 years, or greater than 70 years, depending on how the otoliths had been interpreted.
Validation of this species' age was achieved by analysing levels of 14C (radiocarbon) in otolith cores from fish of a variety of estimated ages. Atmospheric testing of nuclear bombs in the Pacific resulted in an abrupt increase in atmospheric radiocarbon from the late 1950s, and this was incorporated into corals, fish, and other organisms living at the time. There was good agreement between the otolith core 14C measurements for analysed bluenose and their estimated birth years based on the 'abundant narrow' otolith zone interpretation method. Hence, the narrow bands are formed annually and bluenose live to over 70 years old.
Fish ageing (including the development of initial growth curves and their validation, and the routine ageing to provide information for resource assessments) will continue to be an important component of fisheries research. Having comprehensive and accurate growth information is a key factor in successful management of fishery resources.
For further information:
Armiger, H.; Hartill, B.; Rush, N.; Vaughan, M.; Smith, M.; Buckthought, D. (2009). Length and age compositions of recreational landings of kahawai in KAH 1 in January to April 2008 and KAH 8 in January to April 2007. New Zealand Fisheries Assessment Report 2009/36. 40 p.
Campana, S.E. 2001. Accuracy, precision and quality control in age determination, including a review of the use and abuse of age validation methods. Journal of Fish Biology 59: 197–242.
Francis, M.P., Ó Maolagáin, C.; Stevens, D. (2001). Age, growth, and sexual maturity of two New Zealand endemic skates, Dipturus nasutus and D. innominatus. New Zealand J. Mar. Freshwat. Res. 35(4):831-842.
Francis, R.I.C.C.; Paul, L.J.; Mulligan, K.P. 1992. Ageing of adult snapper (Pagrus auratus) from otolith annual ring counts: Validation by tagging and oxytetracycline injection. Australian Journal of Marine and Freshwater Research 43: 1069–1089.
Horn, P.L.; Sullivan, K.J. 1996. Validated aging methodology using otoliths, and growth parameters for hoki (Macruronus novaezelandiae) in New Zealand waters. New Zealand Journal of Marine and Freshwater Research 30: 161–174.
Horn, P.L.; Neil, H.L.; Paul, L.J.; Marriott, P. (2010). Age validation and growth of bluenose (Hyperoglyphe antarctica) using the bomb chronometer method of radiocarbon ageing. Journal of Fish Biology 77: 1552–1563.
Paul, L.J. 1992. Age and growth studies of New Zealand marine fishes, 1921–90: A review and bibliography. Australian Journal of Marine and Freshwater Research 43: 879–912.
Tracey, D.M.; Horn, P.L. 1999. Background and review of ageing orange roughy (Hoplostethus atlanticus, Trachichthyidae) from New Zealand and elsewhere. New Zealand Journal of Marine and Freshwater Research 33: 67–86.