The seaweed known colloquially as nori in Japanese - used for making sushi - or karengo in Maori has been reclassified by an international team of scientists including NIWA's Dr Wendy Nelson.
Overview
Known more scientifically as 'Bangiales', this set of closely-related red algae, or seaweeds, is an ancient order, having survived almost unchanged for over a billion years. In fact, there is a fossil from arctic Canada having been aged at 1,200 million years that looks just like modern species!
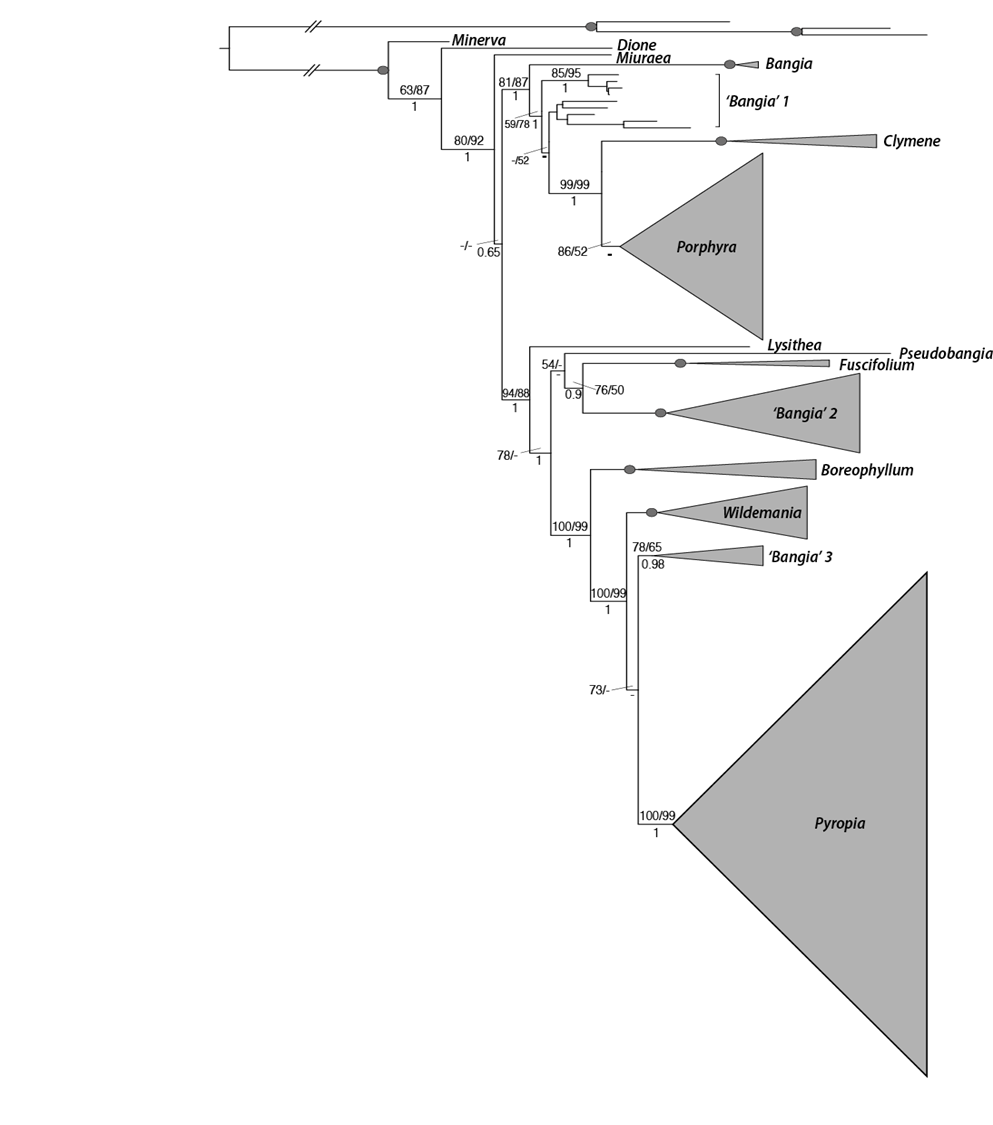
Up until recently, the order has been split into two genera: Bangia (filaments) and Porphyra (blade–like plants). Both of these two genera are found in the intertidal and subtidal areas of the world's oceans.
A 15 year project which reexamined the seaweeds' taxonomy by looking at their genetics has demonstrated that the genetic data does not reflect this difference. As a result, a multi-generic classification was developed for Bangiales:
- filamentous species are now classified as species of Bangia, Dione, Minerva, and Pseudobangia
- Porphyra has now been joined by additional genera: Boreophyllum, Clymene,Fuscifolium, Lysithea, Miuraea, Pyropia, and Wildemania
- additional genera are likely to emerge with further sampling.
In New Zealand we now realise that there are more than 35 different species of karengo, belonging to four different genera, and many of these species still do not have scientific names.
Why is this reclassification important?
The reclassification of Bangiales is important for two primary reasons:
- This will allow producers to improve their breeding programmes, allowing better accuracy and understanding of breeding successes and failures. For example, in the past, people have tried to cross species that we now know belong to different genera. The nori industry is worth over USD 1.4 billion a year, so such understanding is important.
- It is also important for our understanding of evolution and biodiversity.
This research was funded by the Ministry of Business, Innovation & Employment.
Life history
Bangiales has a very complicated life history (we don't call it a 'life cycle' as it's not necessarily a neat cycle!). However, given that related organisms have remained almost unchanged morphologically for over a billion years – and survived the events that caused the KT boundary, unlike many other organisms (including the dinosaurs) at the time – the complexity of the life history may not be surprising, apparently allowing it to survive through almost any sort of environmental conditions.
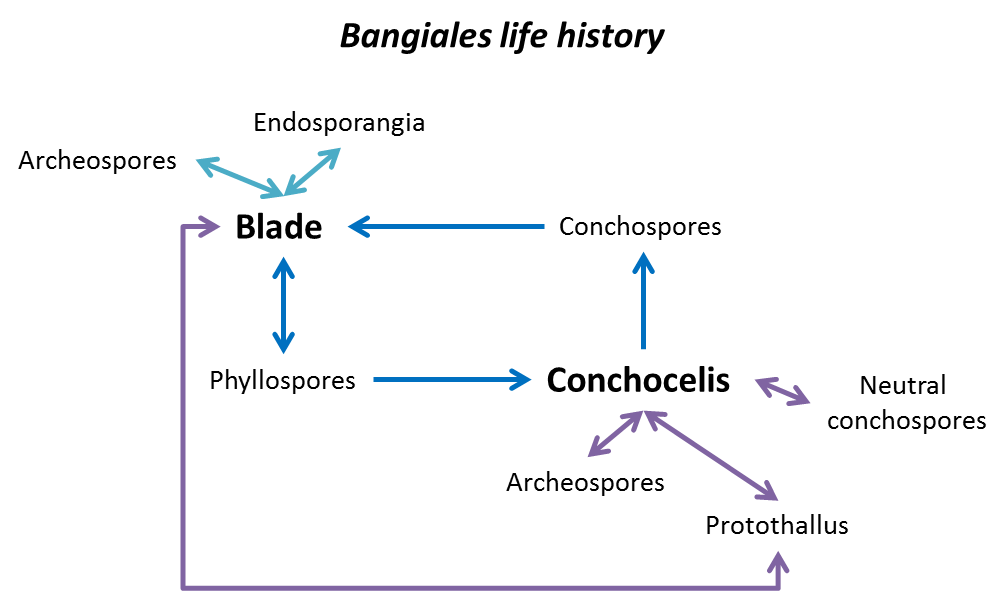
In general, the life history of Bangiales is split into two primary phases: blades and filaments.
Blades
Blades and filamentous thalli are the sexual stage of the life history and produce a range of spores as well as gametes previously assigned to the genus Porphyra, but now also assigned to the genera Clymene or Lysithea.
Blades are only one cell thick and are usually found in the intertidal zone. Filaments range from being very thin and one cell wide, to being broad and made of a number of cell layers. Blades grow underwater.
Until fertilised, all cells in the blades or filaments are 'haploid', which means they have only one copy of their chromosomes. However, the spores produced by blades are generally diploid, and come in various forms:
Phyllospores are single spores, released from their sporangia, or 'packets', and they are produced in a number of ways, the most common of which is the fertilisation of female (haploid) cells by male (haploid) cells called 'spermatia' (we don't call them sperm, as they can't swim). Both the spermatia and the fertilised female cells (or zygotospores) are generally arranged in different patterns of three-dimensional packets – it is the number of cells in these packets, and their arrangement in a blade, which scientists look at first to determine species. These spores are released and form the other primary phase of bangiales: conchocelis (filaments).
Blades also produce other sorts of spores, including archeo- and endospores.
Archeospores (haploid) are formed when a cell on the edge of a blade (still haploid) becomes round, releases itself from the blade, and floats off to form a new blade.
Endospores, described by NIWA scientist Wendy Nelson, are made up of an unformed, 'lumpen' mass of haploid spores which either:
- release themselves to grow into new blades
- self-organise and form into a new blade. This is a process we still don't understand and one which is of great interest to developmental biology.
Conchocelis
The conchocelis phase is the alternate microscopic stage of members of the Bangiales. This phase is so called because the filaments were first found often growing in shells.
As with the blade phase, there are a number of different forms of the conchocelis phase.
Conchosporangia are 'strings' of cells, each of which will release a conchospore. Conchospores will generally go on to form a new blade, going from diploid to haploid in the process (see the Did you know! section below).
Neutral, diploid, conchospores will recycle, becoming new filaments.
Archeospores (diploid), like the archeospores of the blade phase, release themselves from the conchocelis, and form new conchocelis.
Finally, a protothallus, or aggregation of (diploid) filament spores, grows into a blade, through a process we do not yet understand.
Did you know
Blades can be chimeras – that is, they can have different 'flavours' of genes!
Spores are diploid – two sets of chromosomes - so how do the cells which make up blades become haploid (remember: one set of chromosomes per cell)? Bangiales makes use of meiosis to solve this problem.
Meiosis
Meiosis is a special form of cell division used by eukaryotes to make sex cells (such as eggs and sperm, and also blade cells in Bangiales). Meiosis consists of two phases: in the first, a diploid cell doubles its chromosomes and then splits in half, into two 'daughter' cells which are each still diploid, although some recombination has taken place to ensure that the daughter cells are not clones. In the second phases, these two daughter cells then split into two (each) haploid sex cells.
Bangiales, blades and meiosis
The end result of a diploid cell undergoing meiosis is a tetrad of haploid cells, each containing slightly different alleles (gene 'types') of the same genes. The new blade then develops outward from this tetrad, so that sometimes, cells in different parts of the blade are expressing different alleles of a gene: we call this sort of organism a chimera. Experiments in Japan, to demonstrate the chimera quality of blades, have been able to grow them with stripes, for example!